To maintain cellular energy homeostasis mitochondria have to adapt to changes in substrate availability and metabolic demands by modulating oxidative phosphorylation (OXPHOS). Reversible phosphorylation of mitochondrial proteins is an important player in OXPHOS modulation, but only recently the molecular mechanisms have started to emerge. Protein kinase A (PKA) modulates the mitochondrial respiratory chain, and cytochrome oxidase (COX), the terminal enzyme of the electron transfer chain (ETC), is one of the crucial targets of PKA-mediated phosphorylation. COX phosphorylation by PKA is regulated through a metabolic-sensing signaling pathway in the mitochondrial matrix, involving mitochondrial soluble adenylyl cyclase (sAC), which generates cAMP and activates PKA in response to mitochondrial CO2 generated in the Krebs cycle.
COX is one of the three ETC pumps that translocate protons across the inner mitochondrial membrane, creating the proton electrochemical gradient used by the ATP synthase to generate ATP. In living cells, COX is a pacemaker of ETC function, and its activity is modulated by phosphorylation of its subunits and by allosteric ATP inhibition . However, the molecular mechanisms underlying COX regulation in response to changes of substrate availability have been so far completely elusive.
The nuclear-encoded subunits of COX have tissue specific isoforms regulated during development . In particular, subunit IV (COXIV) has two isoforms, COXIV-1 and COXIV-2, which evolutionarily diverged by a gene duplication event about 320My ago. COXIV-1 is expressed ubiquitous and in fetal tissues, while COXIV-2 is highly expressed in adult lung and only at low levels in brain and heart.
We have identified the phosphorylation of a specific amino acid residue in the matrix loop of COXIV-1 as the mechanism for PKA-dependent regulation of COX activity. We show by computational means that this residue is directly involved in binding ATP at the allosteric site, and that its phosphorylation prevents the interaction with the nucleotide.
Molecular Dynamics simulations of membrane embedded dimeric COX
Dimeric WT and mutant COX (COXIV-1 S58A, S58D, and S58phospho) were simulated in the membrane by molecular dynamics (MD) for 1.2ns. A superposition of the most prevalent conformations of the matrix domain of WT and mutant COXIV-1 did not show major structural changes. Thus, the biochemical effects of the S58A and S58D mutations reflect the functional role played by S58 in the regulation of COX activity.
ATP binding site in the matrix domain of COXIV-1
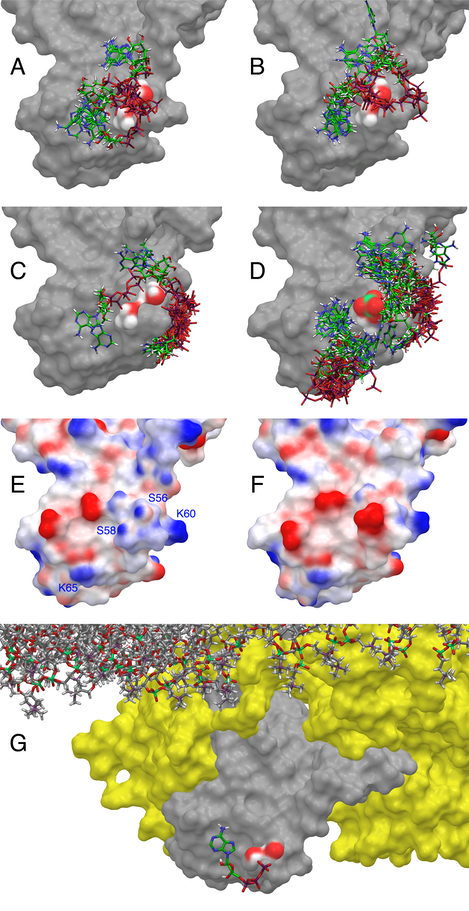
At high ATP:ADP ratios, ATP binds COXIV and other enzyme subunits and reversibly inhibits COX activity . Two different ATP binding sites have been proposed in COXIV, one facing the matrix and regulated by intramitochondrial ATP and a second one facing the intermembrane space and regulated by cytosolic ATP. We hypothesized that phosphorylation of COXIV-1 S58 might regulate the allosteric inhibition of COX by matrix ATP. The binding site for ATP in the matrix domain of COXIV-1 was identified using a combination of Induced Fit Docking (IFD) and QM-Polarized Ligand Docking (QPLD). Five clusters of conformations of the matrix domain of COXIV-1were identified in the MD simulations of WT and S58phospho COXIV-1, and ranked in decreasing order from 1 to 5 based on the number of frames they contained (clusters with more frames reflect more likely conformations). Charges on the ligand (ATP) and protein were assigned based on a mitochondrial matrix pH of 8.0±0.2. In Cluster 1 from WT COXIV-1 there are two groups of ATP binding poses (Panel A) characterized by an interaction of the g- and/or b-phosphate of ATP with the hydroxyl moieties of the side chains of S56 and S58. In Cluster 2 of WT COXIV-1 (Panel B) there is still significant binding of the ATP phosphate chains at the S56/S58 site, but a minor site near to the positive charge of K60 side chain (Panel E) starts being occupied. This second site becomes fully occupied in the remaining conformations (~20% of all the frames, Panel C). S56 and S58 are situated at the N-terminus of the 1st helix of the matrix domain of COXIV-1, and the dipole moment of the helix provides an effective 0.5-0.7 units of positive charge that stabilizes the binding of the g- and b-phosphates of ATP (Panel E). However, the phosphate group of S58phospho obliterates completely this region of positive electrostatic potential (Panel F), thereby preventing the binding of ATP. This is supported by the failure of the IFD protocol to identify a pose corresponding to the binding of ATP with the g- and/or b-phosphates hydrogen bonded to the side chains of S56 and S58 with any of the frames representative of the 5 different conformational clusters of COXIV-1 S58phospho (Panel D). In this case, other binding sites for ATP are recognized near the side chains of K60 and K65, but with poor IFD scores, representing low affinity sites devoid of functional significance. The pose with the best IFD score was refined with the QPLD protocol (Panel G). The phosphate moiety of ATP wraps around the side chain of S58, while the sugar and base moieties are contained in nearby pockets on the surface of COXIV-1. When COXIV-1 is phosphorylated, the phosphate moiety of S58phospho would replace the g-phosphate of ATP, significantly weakening the binding of ATP to the protein. The refined pose shown in Panel G was used to calculate by a free-energy perturbation (FEP) procedure the relative changes in the binding energy of ATP when S58 is replaced by A or D, or is phosphorylated. DDGbind values for ATP with respect to WT were 0.82±2.9 kcal/mol for S58A, 7.15±3.5 kcal/mol for S58D, and 8.6±3.5 kcal/mol for S58phospho (more positive values = weaker binding), corresponding respectively to a ~4-fold, ~175,000-fold, and ~2,100,000-fold increase in Kd.
Key papers:
Hierarchical structure of respiratory complexes
A Tzagoloff, N Capitanio, M Crivelione, D Gatti, M Nobrega
Structural and organizational aspects of metabolic regulation. Alan R. Liss ...
Cloning and sequence analysis of the structural gene for the bc 1-type Rieske iron-sulfur protein from Thermus thermophilus HB8
DL Gatti, G Tarr, JA Fee, SH Ackerman
Journal of bioenergetics and biomembranes 30 (3), 223-233
Structure
and function of the mitochondrial bc1 complex:
A mutational analysis of the yeast rieske iron-sulfur protein
DL Gatti, SW Meinhardt, T Ohnishi, A Tzagoloff
Journal of molecular biology 205 (2), 421-435
Structure and function of the mitochondrial bc1 complex. Properties of the complex in temperature-sensitive cor1 mutants.
DL Gatti, A Tzagoloff
Journal of Biological Chemistry 265 (35), 21468-21475
Cytochrome oxidase assembly in yeast requires the product of COX11, a homolog of the P. denitrificans protein encoded by ORF3.
A Tzagoloff, N Capitanio, MP Nobrega, D Gatti
The EMBO journal 9 (9), 2759
Protein
phosphorylation and prevention of cytochrome oxidase inhibition by ATP:
coupled mechanisms of energy metabolism regulation
R Acin-Perez, DL Gatti, Y Bai, G Manfredi
Cell metabolism 13 (6), 712-719