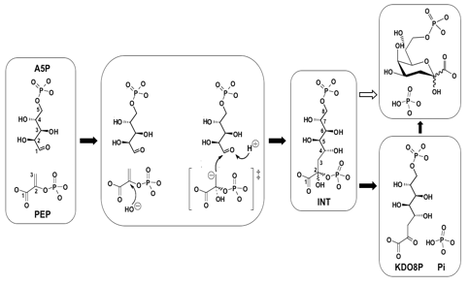
3-Deoxy-D-manno-octulosonate (KDO) is an 8-carbon sugar present in the lipopolysaccharide (LPS) of all Gram negative bacteria. KDO provides a link between lipid A, the membrane embedded moiety of LPS, and the O-antigen, an elongated polysaccharide chain that protrudes from the bacterial outer membrane into the surrounding environment and determines the antigenic specificity of the cell. While the composition of the O-antigen varies between species and also between strains, the inner core region containing KDO is fairly constant among all Gram negative bacteria. 3-Deoxy-D-manno-octulosonate 8-phosphate synthase (KDO8P synthase, EC 4.1.2.16) plays a key role in the biosynthesis of KDO. This enzyme catalyzes the aldol-type condensation of phosphoenolpyruvate (PEP) with arabinose 5-phosphate (A5P) to form KDO8P (precursor to KDO) and inorganic phosphate.
Dephosphorylation of KDO8P to KDO, and synthesis of CMP-KDO (from CTP and KDO) occur prior to insertion of the sugar into LPS. Strains of Salmonella have been isolated with mutations in KDO8P synthase that confer temperature sensitive growth. Such strains fail to synthesize KDO at the non-permissive temperature, which leads to the inhibition of LPS biosynthesis and, as a consequence, to the arrest of cell growth. These studies indicate that KDO8P synthase provides an essential function for bacterial homeostasis, and therefore is a potential drug target.
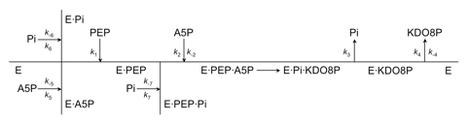
Earlier studies have determined that the reaction of KDO8P synthesis is a sequential process in which the binding of PEP precedes the binding of A5P, and the release of inorganic phosphate precedes the release of KDO8P. The condensation step of the reaction is stereospecific, involving an attack by water (or a hydroxide ion) on the si face of C2PEP and addition of the si face of C3PEP to the re face of C1A5P, to form a doubly phosphorylated non-cyclic intermediate that breaks down into KDO8P and inorganic phosphate (Pi). It has also been established that phosphate release occurs by cleavage of the C-O bond of PEP (9, 14) rather than the P-O bond.
We have reported several structures of both wild-type and mutant metallo-KDO8PS from Aquifex aeolicus in complex with both substrates. This enzyme requires a divalent cation, with different metals (Cd2+, Mn2+, Ni2+, Co2+, Ca2+, Cu2+, Mg2+, Zn2+) providing various levels of activity. Initially, we studied in detail the Cd2+ form of the enzyme because Cd2+ was reported to be the most effective activator of KDO8PS. In subsequent studies we have determined also the structure and kinetic properties of A. aeolicus KDO8PS in complex with Zn2+, Cu2+, and Co2+. In particular, the Cu2+ substituted enzyme allowed the experimental observation of the product KDO8P bound in the active site of the enzyme.
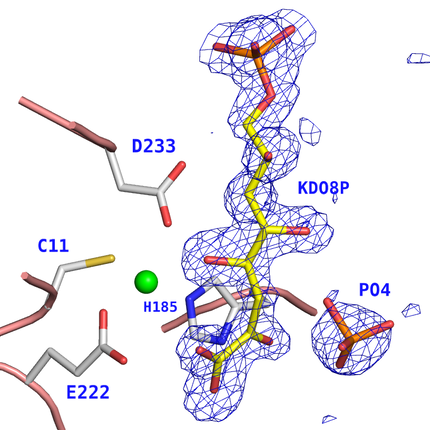
Detail of the active site of Cu2+-KDO8PS with an |FO-FC| sA omit map contoured at 3s around KDO8P (yellow bonds) and Pi (orange bonds).
However, there are two classes of KDO8P synthases characterized respectively by the presence or absence of a metal in the active site. The non-metallo KDO8PS from Escherichia coli and the metallo KDO8PS from Aquifex aeolicus are the best characterized members of each class. All amino acid residues that make important contacts with the substrates are conserved in both enzymes with the exception of Pro-10, Cys-11, Ser-235, and Gln-237 of the A. aeolicus enzyme, which correspond, respectively, to Met-25, Asn-26, Pro-252, and Ala-254 in the E. coli enzyme. Interconversion between the two forms of KDO8P synthases can be achieved by substituting the metal coordinating cysteine of metallo synthases with the corresponding asparagine of non-metallo synthases, and vice versa. We have determined the structural changes elicited by the C11N mutation, and by three combinations of mutations (P10M/C11N, C11N/S235P/Q237A, and P10M/C11N/S235P/Q237A) situated along possible evolutionary paths connecting the A. aeolicus and the E. coli enzyme. All four mutants are not capable of binding metal and lack the structural asymmetry among subunits with regard to substrate binding and conformation of the L7 loop, which is typical of A. aeolicus wild-type KDO8PS, but is absent in the E. coli enzyme. Despite the lack of the active site metal, the mutant enzymes display levels of activity ranging from 46% to 24% of the wild type. With the sole exception of the quadruple mutant, metal loss does not affect the thermal stability of KDO8PS. In all four mutants A5P binding displaces a water molecule located on the si side of PEP. The conformation of the acyclic reaction intermediate, was observed here for the first time in some of the active sites of the triple mutant.
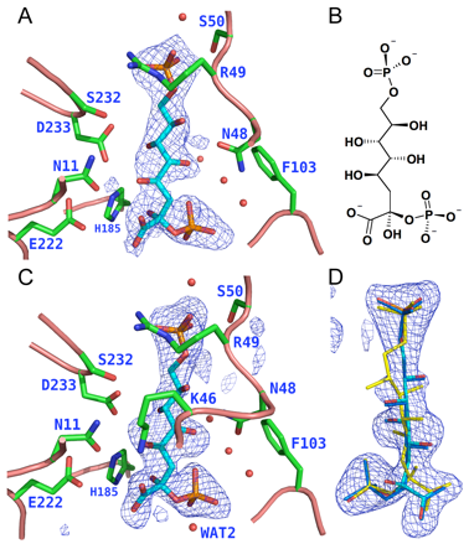
C11N/S235P/Q237A mutant: structure of the reaction intermediate. A. Crystal form C; active site of one of the twelve subunits contained in the asymmetric unit. B. Anticipated chemical structure of INT if water attacks the si side of C2PEP or C2OXO. C. Crystal form D; active site of one of the two subunits contained in the asymmetric unit. D. Detail of the active site shown in C, but rotated by approximately 180°around the vertical axis, showing three alternative interpretations of the electron density: INT produced by water attack from the si side (cyan) or the re side (yellow), and OXO (dark blue). In panels A and C an |FO-FC| sA omit electron density map is contoured at 3s around INT (cyan bonds) produced by water attack from the si side (as in panel B), shown here as the best fit to the electron density.
The direct visualization of this intermediate by X-ray crystallography confirms earlier mechanistic models of KDO8P synthesis. In particular, the configuration of the C2 chiral center of the intermediate supports a model of the reaction in non-metallo KDO8PS, in which water attacks an oxocarbenium ion or PEP from the si side of C2.
Key papers:
Structure and mechanism of 3-deoxy-D-manno-octulosonate 8-phosphate synthase
S Radaev, P Dastidar, M Patel, RW Woodard, DL Gatti
Journal of Biological Chemistry 275 (13), 9476-9484
Substrate and metal complexes of
3-deoxy-D-manno-octulosonate-8-phosphate synthase from Aquifex aeolicus
at 1.9-A resolution. Implications for the condensation
HS Duewel, S Radaev, J Wang, RW Woodard, DL Gatti
Journal of Biological Chemistry 276 (11), 8393-8402
Structure-based design of novel inhibitors of 3-deoxy-D-manno-octulosonate 8-phosphate synthase
X Xu, J Wang, C Grison, S Petek, P Coutrot, M Birck, R Woodard, D Gatti
Drug design and discovery 18 (2-3), 91-99
The catalytic and conformational cycle of Aquifex aeolicus KDO8P synthase: role of the L7 loop
X Xu, F Kona, J Wang, J Lu, T Stemmler, DL Gatti
Biochemistry 44 (37), 12434-12444
Structural
and mechanistic changes along an engineered path from metallo to
nonmetallo 3-deoxy-D-manno-octulosonate 8-phosphate synthases
F Kona, X Xu, P Martin, P Kuzmic, DL Gatti
Biochemistry 46 (15), 4532-4544
Electronic
Structure of the Metal Center in the Cd2+, Zn2+, and Cu2+ Substituted
Forms of KDO8P Synthase: Implications for Catalysis
F Kona, P Tao, P Martin, X Xu, DL Gatti
Biochemistry 48 (16), 3610-3630