This work is conducted in collaboration with Prof. Sharon H. Ackerman.
H+-ATP synthases (F1FO complex) are found in the inner membrane of mitochondria, the thylakoid membrane of chloroplasts, and the plasma membrane of bacteria, where they catalyze the synthesis of ATP from ADP and Pi using the energy of an electrochemical transmembrane proton gradient. These are multimeric proteins with a molecular mass of approximately 550 kDa, whose structure is typically described in terms of two sectors that can be separated from one another under non-denaturing conditions: a hydrophobic domain (FO) that contains the proton channel and a hydrophilic moiety (F1). F1 consists of three a subunits and three b subunits arranged, in alternation, in a hexameric ring that surrounds a rod-like element comprised of a single copy each of g, d, and e subunits. Three of the ab interfaces contain a catalytic site (CS), and three contain a non-catalytic site (NCS). The biogenesis of mitochondrial ATP synthase is a sophisticated event in which the products of two independent genomes associate step-wise in intermediate complexes en route to forming the final structure. Two proteins, Atp11p and Atp12p, were originally identified in Saccharomyces cerevisiae as specialized chaperones required for the assembly of the F1 component of the mitochondrial ATP synthase. It is now established that Atp11p and Atp12p each define a unique protein family (PSSM-Id 26756 and PSSM-Id 14474, respectively), and are required for the assembly of mitochondrial ATP synthase in all eukaryotes. This fact is underscored by the recent report that a genetic lesion of the human locus ATPAF2 encoding Atp12p was responsible for the death of a 14-month old infant. However, Atp11p is not present in prokaryotes, and Atp12p is present only in Proteobacteria, giving further credence to the popular view that mitochondria originated from an a-Proteobacterium ancestor.
S. cerevisiae atp11 or atp12 mutants fail to assemble F1 and are therefore incapable of respiration and growth on a non-fermentable carbon source (e.g., ethanol/glycerol or EG plates). Instead these mutants accumulate aggregated a and b subunits in the mitochondrial matrix as inclusion bodies, an observation that led initially to the hypothesis that Atp11p and Atp12p prevent non-productive protein-protein interactions during the assembly of F1 as a soluble unit, prior to its association with FO in the membrane. More recent studies in yeast have shown that Atp11p binds to the F1 b subunit, and that Atp12p binds to the F1 a subunit. While the structure of F1 from different sources is known, the nature of the interactions between the F1 subunits and Atp11p and Atp12p has remained elusive due to the lack of structural information about these chaperones. We have determined the structures of Atp11p from Candida glabrata and of Atp12p from Paracoccus denitrificans. Crystals of these proteins were obtained after screening several homologous proteins including those from Homo sapiens, Mus musculus, S. cerevisiae, Candida albicans, Kluveromyces lactis, Rhodobacter capulatus, and Arabidopsis thaliana. Atp11p from C. glabrata and Atp12p from P. denitrificans are 79.2% and 44.2% similar, and 56.3% and 23.4% identical to the own proteins in S. cerevisiae (Fig. S1), and when expressed from an episomal plasmid rescue S. cerevisiae atp11 or atp12 mutants, restoring wild-type growth on EG plates (data not shown). Thus, the essential features of these important proteins are maintained across the prokaryotic and eukaryotic kingdoms.
Structure of Atp11p.
The crystal structure of C. glabrata Atp11p was determined at the resolution of 1.4 Å. The first 93 residues (out of 298) are not visible, although the full-length protein produced in E. coli can be recovered from the crystals, and an empty cavity in the density map is large enough to harbor the disordered domain. Residues 94-298 correspond to the part of S. cerevisiae Atp11p that was shown by deletion studies to retain full chaperone activity. A small region of this domain (residues 163-176), which is poorly conserved or absent in Atp11p proteins from different sources, is also disordered. The overall fold recalls what might be called an a/b “taco” (an incomplete antiparallel b-barrel whose concave face is filled by two a-helices) flanked by two helical domains.
Structure of Atp12p
The structure of P. denitrificans Atp12p was determined at the resolution of 1.0 Å. In the asymmetric unit of crystal form I there are two chains of Atp12p related by a local 2-fold axis, but they are loosely packed and unlikely to represent a functional dimer. A single chain occupies the asymmetric unit of crystal form II, with no symmetry neighbors in the unit cell in a position suitable to form a dimer with a large buried surface. These data indicate that P. denitrificans ATP12p is a monomer in the crystalline environment, and agree with earlier sedimentation, cross-linking and two-hybrid studies of yeast and human Atp12p. The overall shape of Atp12p resembles that of a boxing glove, with the wrist and palm sections provided by two domains encompassing residues 3-83 (N-domain) and 84-238 (C-domain). The two-domain structure of the P. denitrificans Atp12p explains earlier observations with homologous forms of this protein. For example, the GdnHCl denaturation curve of the H. sapiens protein is bimodal. Deletion of the sequence of S. cerevisiae Atp12p corresponding to the first 37 residues of the P. denitrificans sequence produces a protein that still supports ~80% of the wild-type level of F1 assembly in vivo. A larger deletion of the first 77 residues produces a temperature sensitive protein that supports only ~14% of the wild-type level of F1 assembly when yeast are cultured at 30°C, but up to ~60% at 23°C. Substitution of Glu289 in the C-domain of S. cerevisiae Atp12p (corresponding to Asp202 in P. denitrificans )(GREEN surface in the figure above) with Lys eliminates F1 assembly in vivo without decreasing the amount of protein detected in isolated mitochondria, and thus appears to affect directly the activity of the chaperone. These studies indicate that only the larger C-domain of Atp12p is necessary for activity, while the N-domain is necessary for stability.
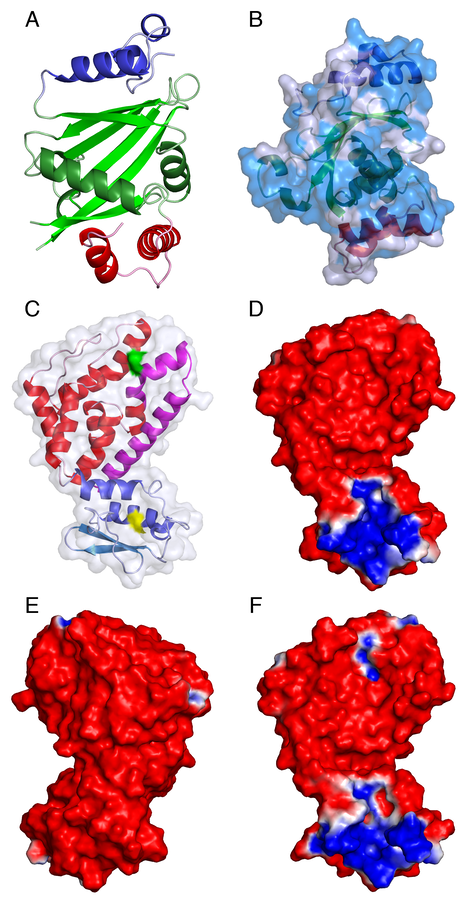
Structures of C. glabrata Atp11p and P. denitrificans Atp12p. A. The N-terminal helical domain (residues 94-129), the central a/b “taco” (residues 130-262), and the C-terminal helical domain (residues 263-298) of C. glabrata Atp11p are shown as a cartoon colored in blue, green and red, respectively. Loops are shown as lighter colors. B. The solvent accessible surface of Atp11p, rotated by approximately 90 degrees with respect to panel A,is colored in transparent light blue with the regions of identity with the S. cerevisiae protein shown in a darker shade: the internal structure is shown as a cartoon colored as in panel A. C. Characteristic “boxing glove” shape of P. denitrificans Atp12p. The N-terminal “wrist” domain (residues 3-83), and the C-terminal “hand” domain (residues 84-238) are shown as a cartoon colored in shades of blue/light-blue and red/magenta, respectively. The first 37 residues of the P. denitrificans protein, corresponding to a deletion of the S. cerevisiae protein that severely affects stability, are shown in light blue. This region corresponds approximately to an N-terminal b-hairpin. The last 42 residues of the P. denitrificans protein, corresponding to a deletion of the S. cerevisiae protein that completely eliminates the chaperone activity, are shown in magenta. The surfaces of Trp57 and Asp202, corresponding to Trp103 and Glu289 of the yeast protein, are shown in yellow and green, respectively: mutations of these residues also inactivate the protein. D-F. The palm (panel D) and dorso (panel E) faces of wild-type Atp12p, and the palm face of D202K (panel F) Atp12p from P. denitrificans are shown colored according to the electrostatic potentials (-1kT/e, red; 1kT/e, blue) at their solvent accessible surface.
Physical forces involved in F1 assembly.
Earlier models on the role of Atp11p and Atp12p were based on the hypothesis that the regions of F1 a and b subunits most likely to promote aggregation are hydrophobic patches sequestered at the subunit interfaces in the a3b3 structure, and that Atp11p and Atp12p mimic the aand bsubunit, temporarily masking these hydrophobic regions. F1 assembly would occur by exchange of Atp11p for a and Atp12p for b, and sequential addition of individual a and b subunits until a hexamer is formed. However, side-by-side analysis of the structures of both F1 and the two chaperones shows no similarity of shape between Atp11p and a and Atp12p and b, and no dramatic difference in the extent and distribution of hydrophobic patches between the regions of F1 a and b subunits exposed to solvent, and those buried at the subunits interface. Furthermore, hydrophobic areas account for less than 50% of the total contact surface between these subunits. Thus, other forces, besides hydrophobic interactions, play an important role in F1 assembly. When electrostatic potentials are computed separately for each individual subunit of F1, the regions of the a subunits that contact the b subunits appear mostly positively charged, and the corresponding regions of the b subunits appear mostly negatively charged. Remarkably, the surface potentials of F1 b and a subunits computed in this way complement well the surface potentials of the Atp11p dimer and of Atp12p. These observations suggest that electrostatic attraction plays a major role in the interaction of F1 aand bwith their cognate chaperones and in the formation of the correct interfaces in F1.
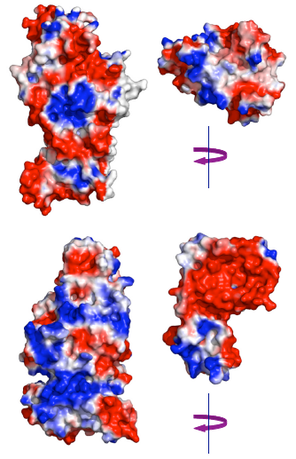
UPPER: electrostatic potentials of the CS interface of yeast F1 bD subunit (PDB entry 2HLD) and of the Atp11p surface expected to interact with bD. LOWER: electrostatic potentials of the CS interface of yeast F1 aA subunit and of the “palm” face of Atp12p expected to interact with aA. The interactions can be visualized by rotating Atp11p over bD and Atp12p over aA around a vertical 2-fold axis. F1 subunits and chaperons are colored according to the electrostatic potentials (-1kT/e, red; 1kT/e, blue) at their solvent accessible surface.
Intermediates in F1 assembly.
The fact that Atp11p does not resemble a and Atp12p does not resemble b is inconsistent with assembly mechanisms that require some degree of molecular mimicry between Atp11p and a, and between Atp12p and b. In these models, the chaperone is released (e.g from b) in exchange for the incoming partner subunit (e.g. a), and F1 is built sequentially with consecutive additions of a and b subunits until the hexamer is formed. The likelihood of F1 assembly occurring through a series of progressively larger intermediates was studied in yeast cells in which the nuclear gene encoding the F1 g subunit was disrupted. Dg cells are incapable of respiration and growth on a non-fermentable carbon source (e.g., EG plates). Despite normal amount of a and b being present in soluble mitochondrial extract, there is little or no hexameric F1 and no F1 (e.g. oligomycin sensitive) ATPase activity. Surprisingly, in this mutant a and b subunits do not precipitate in the mitochondrial matrix as inclusion bodies, but rather accumulate as soluble a:b dimers, suggesting that these dimers, which do not carry any of the chaperones, are the main intermediate along the path to F1 assembly. This result also indicates that the g subunit is necessary for the final assembly of free a:b dimers into mature F1. As Atp11p can be co-precipitated as a stable complex with the b subunit, and Atp12 with the a subunit, formation of a transient quaternary complex Atp11p:b:a:Atp12, as an assembly intermediate, must be rapidly followed by release of the chaperones leaving a soluble a:b dimer. We have used the automatic docking routine of the program Zdock to identify potential complexes of Atp11p with b and Atp12p with a that would best reflect both the surface and electrostatic complementarity of these proteins. Based on Zdock performance in the CAPRI (Critical Assessment of Prediction of Interactions) survey, there is a high probability that the correct complex is found among the top 10 scoring solutions identified by this program. However, there is no certainty that the best scoring solution is also the correct one, and in most cases additional information is necessary to identify the real complex. In the case of Atp11p and Atp12p, the correct complexes must include interaction of the chaperone with the NBD domain of a and b, as determined in earlier experimental studies; in addition Atp12p must bind to a through its “palm” face. The second best scoring solutions for the docking of Atp11p with b, and the third best solution for the docking of Atp12p with a fulfill these requirements. The surface potential of the Atp11p:b and Atp12p:a complexes show significant charge complementarity, the former being mostly positive, and the latter mostly negative (Figure below A,B). Furthermore, when the complexes are brought together by pairing a and bat their NCS interface, there is no steric clash between the two chaperones (Figure below C,D). In particular, the position of the Atp11p dimer is such that also the N-terminal disordered domain (first 93 residues) could be easily accommodated without displacing Atp12p.
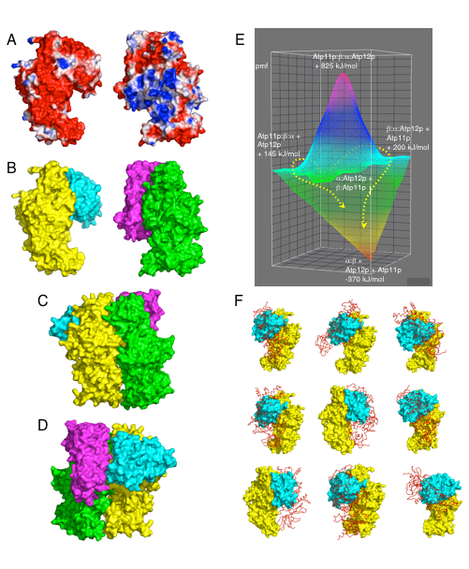
Model of F1 assembly. A. NCS interfaces of the yeast F1 bD subunit with bound Atp11p (left), and of the aA subunit with bound Atp12p (right) colored according to the electrostatic potentials (-1kT/e, red; 1kT/e, blue) at their solvent accessible surface. B. Same complexes as in panel A, but colored by chain: bD, yellow; Atp11p, cyan; aA, green; Atp12p, magenta. C. Quaternary complex Atp11p:bD:aA:Atp12p, showing the surface of bD and aA that is facing outward in F1. D. Same complex, but showing the surface of bD and aA that is facing inward in F1. E. Potential energy surface of the reaction leading to a:b + Atp11p + Atp12p from Atp11p:b + a:Atp12p (taken as the reference state). Dashed yellow arrows show two nearly isoenergetic reaction coordinates. Potential of mean force (pmf) differences (kJ/mol) were calculated with respect to the reference state. F. Atp11p (cyan surface) bound to b (yellow surface) as shown in Panel B (left) prevents 9 of the top 10 docking modes of a second b (red ribbon) to the first one. Each panel shows significant overlap between one of the two b subunits and Atp11p.
At this time it is not clear what might trigger the release of the chaperones from the transient quaternary complex. Attempts to identify small molecular weight cofactors modulating the binding of Atp11p and Atp12p to the F1 subunits have not been successful. Thus, it is conceivable that binding of the Atp11p:b to the Atp12p:a complex might produce some conformational change that decreases the affinity of the chaperones for their target proteins. Another possibility is that the coming together of a and b at their NCS interface changes the surface electrostatic potential of the dimer with respect to the individual subunits, and that this change might be sufficient to detach the chaperones, whose binding was seen to have a significant electrostatic component. With regard to the final steps leading to the formation of mature F1, the aforementioned studies with the Dg yeast strain clearly indicate that, at least in eukaryotic cells, the g subunit is necessary for the final gathering of three separate a:b dimers. It is of note that the g subunit has only been observed to interact with the CS interfaces of a and b, and that in assembled F1 ATP hydrolysis at the CS produces rotation of the g subunit to the next CS. Thus, it is tempting to speculate that upon formation of an initial complex of two a:b dimers, containing the g subunit at the CS interface, unisite ATP hydrolysis would produce a rotation of this subunit to the next CS and thus favor the addition of the final a:b dimer. A sequential model for the complete assembly of F1 is shown below.
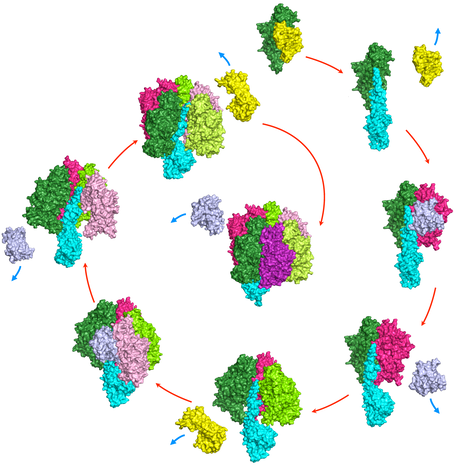
Chaperone-mediated assembly of F1 ATPase. Assembly starts with F1-g displacing Atp12p (yellow) from the Atp12p:a heterodimer exposing the a NCS surface to which Atp11p:b binds. Displacement of Atp11p (violet) exposes the b CS surface to which a second Atp12p:a binds. The cycle continues with consecutive additions of Atp11p:b and Atp12p:a and concurrent release of chaperones, until F1 is assembled.
Role of Atp11p and Atp12p.
Atp11p and Atp12p are present in the mitochondrial matrix in minute amounts, comparable to those of unassembled a and b subunits. In the absence of one of these subunits, as F1 cannot be assembled, the concentration of the other subunit exceeds that of the cognate chaperone and the subunit aggregates. In S. cerevisiae strains harboring a desruption of the nuclear gene encoding either the a or the b subunit, insoluble aggregates are formed even in the presence of normal amounts of functioning Atp11p and Atp12p. In silico docking of a with a and b with b suggests that these subunits can form stable complexes in a large number of ways. ATP11p bound to b and Atp12p bound to a (as shown above) prevent the formation of 9 of the top 10 binding modes of b with b and of a with a. Thus, the predicted structures of Atp11p:b and Atp12p:a complexes suggest that these chaperones are required in F1 assembly to prevent the formation of non-physiological a:a and b:b complexes.
At this time it is not completely clear why Atp11p and Atp12p are not necessary in some bacteria. The fact that Atp12p, but not Atp11p, is present in P. denitrificans is possibly related to the different solubility properties of the F1 a and b subunits in this microorganism. For example, when P. denitrificans F1 a is produced in E. coli it forms insoluble aggregates unless co-expressed with Atp12p. In contrast, P. denitrificans F1 b is produced in E. coli at very high level in soluble form (not shown). On this basis, it is reasonable to suggest that Atp11p and Atp12p are necessary when the electrostatic properties of F1 a and b do not favor the formation of ordered hexamers over other more random assemblies. In humans, only one fatal syndrome has been attributed to a defect in an F1 chaperone. However, intermediate phenotypes with partially impaired assembly of F1 are likely to exist, in which a metabolic defect will appear only under stress. Other unrelated pathologic conditions are also characterized by the formation of non-physiological aggregates. For example, it has been suggested that coulombic interactions play a key role in the formation of amyloid Ab fibrils, which is the central event in the development of Alzheimer disease, and at least two distinct chaperones have been implicated in preventing the formation of these fibrils. Thus, the mechanism by which Atp11p and Atp12p prevent the chaotic aggregation of F1 subunits might be a useful model to understand the function of other dedicated chaperones, which also appear to act primarily by masking electrostatic interactions.
Key papers:
Labeling of individual amino acid residues in the membrane‐embedded F0 part of the F1 F0 ATP synthase from Neurospora crassa
J HOPPE, D GATTI, H WEBER, W Sebald European Journal of Biochemistry 155 (2), 259-264
The α-subunit of the mitochondrial F1 ATPase interacts directly with the assembly factor Atp12p
ZG Wang, D Sheluho, DL Gatti, SH AckermanThe EMBO journal 19 (7), 1486-1493
The Molecular Chaperone, Atp12p, from Homo sapiens: in vitro studies with purified wild-type and mutant (E240K) proteins.
A Hinton, DL Gatti, SH Ackerman
Journal of Biological Chemistry 279 (10), 9016-9022
Chaperones of F1-ATPase
A Ludlam, J Brunzelle, T Pribyl, X Xu, DL Gatti, SH Ackerman
Journal of Biological Chemistry 284 (25), 17138-17146
Defining the pathogenesis of the human Atp12p W94R mutation using a Saccharomyces cerevisiae yeast model
A Meulemans, S Seneca, T Pribyl, J Smet, V Alderweirldt, A Waeytens, W ...
Journal of Biological Chemistry 285 (6), 4099-4109